A new article about facilitation cascades has been published by Christine Angelini and Brian Silliman from the University of Florida. It is such an interesting contribution to our understanding of epiphyte ecology that I'm going to dedicate four blogs to it. This week: a model to explain the application.
To understand and predict species assemblages and interactions in different facilitation cascades we can use the Foundation Species-Biodiversity model:
To understand and predict species assemblages and interactions in different facilitation cascades we can use the Foundation Species-Biodiversity model:
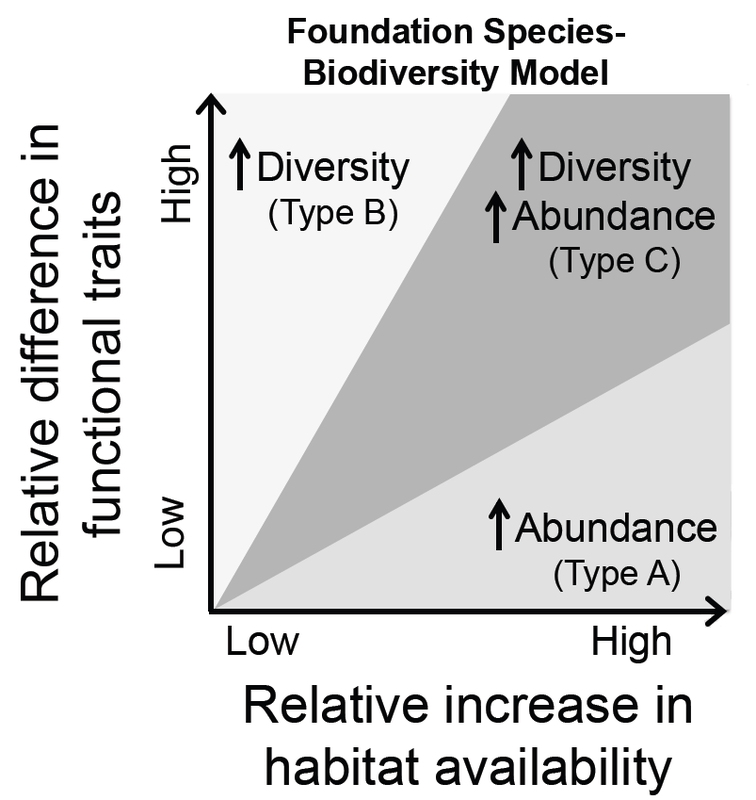
Foundation Species-Biodiversity Model from Angelini and Silliman (In Press).
According to this model, facilitation cascades can be described using three categories based on the difference that secondary foundation species make to the community:
Type A:
Secondary foundation species support more individuals of species that are already in the community (increased abundance) by providing functional traits that are similar to those of the foundation species.
Type B:
Secondary foundation species provide habitat for different individuals (increased diversity) by providing different functional traits than the foundation species.
Type C:
Secondary foundation species support more individuals of the existing species AND individuals of new species (increased abundance AND diversity) through the provision of a range of traits.
Notes: some secondary species may not significantly increase either abundance or diversity, and facilitation cascades can switch if environmental conditions change.
Type A:
Secondary foundation species support more individuals of species that are already in the community (increased abundance) by providing functional traits that are similar to those of the foundation species.
Type B:
Secondary foundation species provide habitat for different individuals (increased diversity) by providing different functional traits than the foundation species.
Type C:
Secondary foundation species support more individuals of the existing species AND individuals of new species (increased abundance AND diversity) through the provision of a range of traits.
Notes: some secondary species may not significantly increase either abundance or diversity, and facilitation cascades can switch if environmental conditions change.

Manatee grass. Photo: Project Noah.
Examples of each facilitation cascade. Simplified from Angelini & Silliman:
Type A:
Foundation species: shoal grass
Secondary foundation species: manatee grass
Manatee grass facilitates an increase in the abundance of resident invertebrates and fish by providing more habitat of a similar nature. In other words, more of the same services.
Type B:
Foundation species: turtle grass
Secondary foundation species: pen shells
Pen shells facilitate increased diversity by providing places for fish to lay eggs. In other words, new and different services.
Type C:
Foundation species: mangroves
Secondary foundation species: oysters
Oysters facilitate both an increase in diversity and abundance. They provide more habitat for species that already inhabit mangrove roots but also provide habitat for species that are oyster-dependant.
Type A:
Foundation species: shoal grass
Secondary foundation species: manatee grass
Manatee grass facilitates an increase in the abundance of resident invertebrates and fish by providing more habitat of a similar nature. In other words, more of the same services.
Type B:
Foundation species: turtle grass
Secondary foundation species: pen shells
Pen shells facilitate increased diversity by providing places for fish to lay eggs. In other words, new and different services.
Type C:
Foundation species: mangroves
Secondary foundation species: oysters
Oysters facilitate both an increase in diversity and abundance. They provide more habitat for species that already inhabit mangrove roots but also provide habitat for species that are oyster-dependant.

Oysters on mangrove roots. Photo: Ji-Elle.
Next week: application of all of this theory to New Zealand epiphytes!






 RSS Feed
RSS Feed
